
Towards a natural system of organisms: Proposal for the domains Archae, Bacteria, and Eucarya
EXTRACTS FROM
Proc. Natl. Acad. Sci. USA
Vol. 87, pp. 4576 - 4579, June 1990
Evolution
CARL R. WOESE, OTTO KANDLER, AND MARK I. WHEELIS
ABSTRACT Molecular structures and sequences are generally more revealing of evolutionary relationships than are classical phenotypes (particularly so among microorganisms). Consequently,the basis for the definition of taxa has progressively shifted form the organismal to the cellular to the molecular level. Molecular comparisons show that life on this planet divides into three primary groupings, commonly known as the eubacteria, the archaebacteria, and the eukaryotes. The three are very dissimilar, the differences that separate them being of a more profound nature than the differences that separate typical kingdoms, such as animals and plants. Unfortunately, neither of the conventionally accepted views of the natural relationships among living systems -- i.e., the five-kingdom taxonomy or the eukaryote-prokaryote dichotomy -- reflects this primary tripartite division of the living world. To remedy this situation we propose that a formal system of be established in which above the level of kingdom there exists a new taxon called a "domain." Life on this planet would then be seen as comprising three domains, the Bacteria, the Archaea, and the Eucarya, each containing two or more kingdoms. (The Eucarya, for example, contains Animalia, Plantae, Fungi, and a number of others yet to be defined.) Although taxonomic structure within the Bacteria and Eucarya is not treated herein, Archaea is formally subdivided into the two kingdoms Euryarchaeota (encompassing the methanogens and their phenotypically diverse relatives) and Crenarchaeota (comprising the relatively light clustering of extremely thermophilic archaebacteria, whose general phenotype appears to resemble most the ancestral phenotype of the Archaea).
Basis for Restructuring
What must be recognised is that the basis for systematics has changed; classical phenotypic criteria are being replaced by molecular criteria. As Zuckerkandl and Pauling (2) made clear many years ago, it is at the level of molecules (particularly molecular sequences) that one really becomes privy to the workings of the evolutionary process. Molecular sequences can reveal evolutionary relationships in a way and to an extent that classical phenotypic criteria, and even molecular functions, cannot; and what is seen only dimly, if at all, at the higher levels of organisation can be seen clearly at the level of molecular structure and sequence. Thus systematics in the future will be based primarily upon the sequences, structure, and relationships of molecules, the classical gross properties of cells and organisms being used largely to confirm and embellish these.
It is only on the molecular level that we see the living world divide into three distinct primary groups. For every well-characterised molecular system there exists a characteristic eubacterial, archaebacterial, and eukaryotic version, which all members of each group share. Ribosomal RNAs provide an excellent example (in part because they have been so thoroughly studied). One structural feature in the small subunit rRNA by which the eubacteria can be distinguished from the archaebacteria and eukaryotes is the hairpin loop lying between positions 300 and 545 (3), which has a side bulge protruding from the stalk of the structure. In all eubacterial cases (over 400 known) the side bulge comprises six nucleotides (of a characteristic composition), and it protrudes from the "upstream" strand of the stalk between the fifth and sixth base pair. In both archaebacteria and eukaryotes, however, the corresponding bulge comprises seven nucleotides (of a different characteristic composition), and it protrudes from the stalk between the sixth and seventh pair (3, 4). The small subunit rRNA of eukaryotes, on the other hand, is readily identified by the region between positions 585 and 655 (E. coli numbering), because both prokaryotic groups exhibit a common characteristic structure here that is never seen in eukaryotes (3, 4). Finally archaebacterial 16S rRNAs are readily identified by the unique structure that they show on the region between positions 180 and 197 or that between positions 405 and 498 (3, 4). Many other examples of group-invariant rRNA characteristics exist;see refs. 1, 3 and 4. [The reader wishing to gain a broader and more detailed appreciation for the molecular definition of the three groups can consult refs. 1, 5 and 6 and the proceedings of the most recent conference on archaebacteria (7).]
Molecular characterisations also reveal that the evolutionary differences among the eubacteria, archaebacteria, and eukaryotes are of a more profound nature than those that distinguish traditional kingdoms, such as animals and plants, from one another. This is most clearly seen in the functions that have evolved early in the cell's history and are basic to its workings. All eubacteria, for example, exhibit nearly the same subunit pattern (in terms of numbers and sizes) in their RNA polymerases; however, this pattern bears little relationship to that seen in either the archaebacteria or the eukaryotes (8). On the other hand, eukaryotes are unique in using three separate RNA polymerase functions (9).
The fossil record indicates that photosynthetic eubacteria (and by inforence, therefore, archaebacteria and possibly eukaryotes) were already in existence 3 - 4 billion years ago (10), so that the evolutionary events that transformed the ancestor common to all life into the individual ancestors of each of the three major groups must have occurred over a relatively short time span early in the planet's history. Both the relatively rapid pace of, as well as the profound changes associated with, this early evolution argue that the universal ancestor was a simpler, more rudimentary entity than the universal ancestors that spawned the three groups (and their descendents) (11).
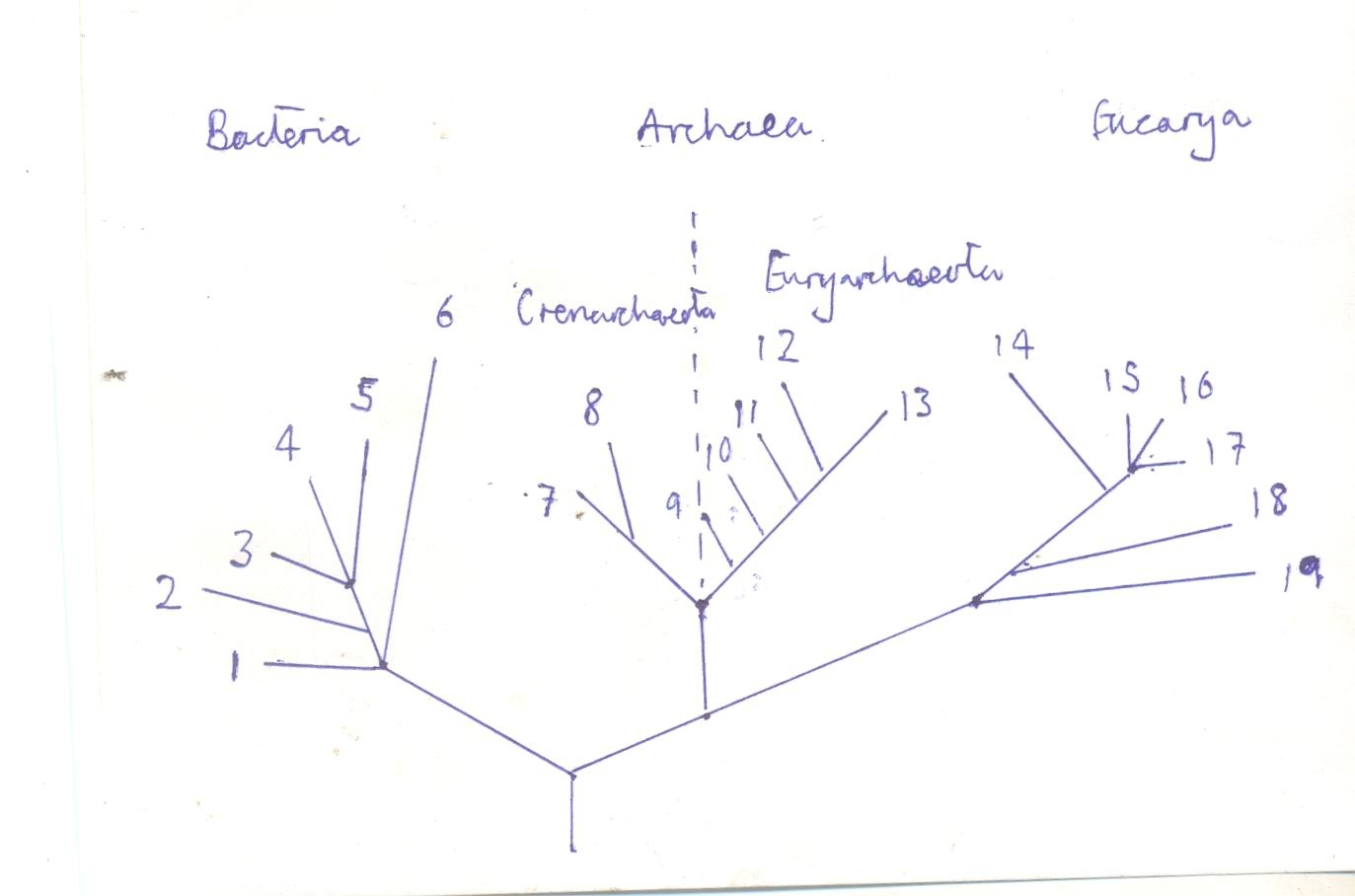
|
| FIG 1. Universal phylogenic tree in rooted form, showing the three domains. Brabching order and branch lengths are based upon rRNA sequence comparisons (and have been taken from figure 4 of ref. *). The position of the root was determined by comparing (the few known) sequences of pairs of paralogous genes that diverged from each other before the primary lineages emerged from their common ancestral condition (*). [This rooting strategy (*) in effect uses the one set of (aboriginally duplicated) genes as an outgroup for the other.] The numbers on the branch tips correspond to the following groups of organisms (*). Bacteria: 1. the Thermotogales; 2. the flavobacteria and relatives; 3. the cyanobacteria; 4. the purple bacteria; 5. the Gram-positive bacteria; and 6. the green nonsulphur bacteria. Archae: the kingdom Crenarchaeota: 7.the genus Pyrodictium and 8. the genusThermoproteus; and the kingdom Euryarchaeota:9. the Thermococcales; 10. the Methanococcales; 11. the Methanobacteriales; 12.the Methanomicrobiales; and 13. the extreme halophiles. Eucarya: 14. the animals; 15.the ciliates; 16. the green plants; 17. the fungi; 18. the flagellates; and 19. the microsporidia. |
| Comprehensive information may be found at Microbial Biorealm - MicrobeWiki |
Fig. 1 is a universal tree, showing the relationships among the primary groups. The root of the tree is seen to separate the eubacteria from the other two primary groups,making the archaebacteria and eukaryotes specific (but distant) relatives. A relationship between archaebacteria and eukaryotes is not overly surprising, for with a few exceptions (the rRNA being one) the archaebacterial versions of molecules resemble their eukaryotic homologs more than their eubacterial ones (9, 12, 13). Ampng the ribosomal proteins there are even cases where the archaebacterial and eukaryotic homologs have no apparent counterpart among the eubacteria (12, 13).
From a systematic perspective the specific relationship between eukaryotes and archaebacteria does not require taxonomic recognition; these two groups are sufficiently dissimilar, and they diverges so early, that little would be gained by defining a taxon that encompasses both. In other words, the archaeobacteria and eukaryotes themselves show the kind of profound molecular differences that distinguish either from the eubacteria.
References
See also: Classification of Organisms
--- LIFE SCIENCES ---
